Переклад Анни Стембковської
Фахова редакція Дмитра Залеського, Ірини Семків
Antonio Alcaro1,2, Stefano Carta3 і Jaak Panksepp4
REVIEW article
Front. Psychol., 01 September 2017 | https://doi.org/10.3389/fpsyg.2017.01424
Психологи зазвичай розглядали «Самість» як об’єкт досвіду, який з’являється тоді, коли індивід сприймає його в полі свідомості. Відповідно до цього погляду, здатність людської психіки до самопрезентації пов’язується з кортиколімбічними процесами навчання, що відбуваються впродовж індивідуального розвитку. З іншого боку, Карл Густав Юнг розглядав Самість як ядро нашої особистості, в її свідомих і несвідомих аспектах, а так само як і у її актуальних і потенційних формах. Згідно з Юнгом, Самість походить від вродженої динамічної структури, що інтегрує значущі потяги нашого «мозку-розуму» та веде як до інстинктивних поведінкових дій, так і до архетипових психологічних переживань. Цікавим є те, що останні нейроетологічні дослідження вказують на те, що наша суб’єктивна ідентичність ґрунтується на архаїчних нейропсихічних процесах, які в людей спільні з іншими тваринами та є складовою їхнього вродженого конституційного репертуару. Дійсно, мозкова діяльність у підкіркових структурах середнього мозку (ПКСС) внутрішньо пов’язана з появою прототипних афективних станів, які не лише гнучко впливають на нашу поведінку, але й будучи джерелом специфічних переживань та настроїв (які є першою формою самоорієнтації у світі) змінюють поле нашої свідомості. Більше того, така афективна динаміка відіграє центральну роль в організації індивідуальності особистості та в еволюції всіх інших (складніших) психологічних функцій. Таким чином, на основі зближення найбільш сучасних наукових досліджень та деяких психологічних інтуїтивних здогадок Юнга, ми маємо намір дослідити тут перший нейроеволюційний прошарок людського розуму, який ми називаємо Афективним Ядром Самості.
«Я давно думав, що, якщо існує якась аналогія між психічними та фізіологічними процесами, то організуюча система мозку мала б лежати в підкірці у стовбурі мозку. Це припущення випливає з міркувань про психологію архетипу [Самості], який має центральне значення та універсальне розповсюдження, що представлене у символах мандали. Причиною, яка підштовхнула мене припустити, що фізіологічне підгрунтя цього архетипу локалізоване у стовбурі мозку, був той психологічний факт, що об’єднуючі властивості цього архетипу є здебільшого афективними, окрім цього, він має організуюче та орієнтувальне значення. Я б припустив, що така підкіркова система може якимось чином відображати характеристику архетипової форми несвідомого». Jung (1958, para. 582).
Jung (1958, para. 582).
У західній філософській і науковій традиції психічне суб’єктивне життя, що виникає внаслідок діяльності високорозвинених і складних неокортикальних когнітивних функцій загалом розглядається, як прерогатива людських істот. Ці погляди підтримуються популярною нейрокогнітивною парадигмою, яка все ще використовується для пояснення локалізації суб’єктивних переживань у вищих лімбічних і кортикальних ділянках мозку (LeDoux, 1996; Rolls, 1999; Kandel, 2005), просуваючи переконання, що свідомість виникає лише зсередини таламокортикальних мереж (Edelman, 1989; Crick, 1994). Відповідно до таких поглядів, психоаналітики та когнітивні психологи вважали, що суб’єктивність набувається під час індивідуального розвитку людини та оходить з інтроєкції деяких, пов’язаних із прив’язаністю робочих моделей, що призводить до свого роду рефлексивного самосприйняття (Stern, 1985; Schore, 1994; Fonagy, 2001).
Переконлива версія такого кортикокогнітивного антропоцентризму нещодавно була перевірена в декількох публікаціях Джозефом Леду (LeDoux, 2015; LeDoux and Brown, 2017). Він розглядає суб’єктивний досвід людини як результат кортикальних процесів вищого порядку, що залучені до когнітивних інтерпретацій нашого буття в світі. Обмеження такого підходу випливає з помилкової думки, що суб’єктивний досвід обов’язково пов’язана з фактом самоусвідомлення цього досвіду, оскільки це можна перевірити, запитавши в людини «що ти відчуваєш?». І навпаки, багато різних дослідників у галузі психології, філософії та нейронауки визнали існування суб’єктивних переживань без самоусвідомлення, що, мабуть, постає лише вторинним еволюційним продуктом нашого психічного життя (James, 1890/1950; Edelman, 1992; Searle, 1992; Damasio, 2010).
Відповідно до емпіричної точки зору James (1890/1950), нейрофілософ Георг Нортхофф визначив «феноменальну мінімальну Самість» як дорефлексивну форму суб’єктивності. Вона включає досвід, який визначається певними (до)свідомими якостями, або «кваліа», та імпліцитним відчуттям приналежності до цього досвіду (Northoff, 2013; Northoff et al., 2014). Перевагою такого визначення є звернення уваги на дві необхідні і достатні характеристики Самості: інтенціональності (наміреності) та свідомої сенситивності (чутливості) (McGinn, 1989; Searle, 1991). Наміреність (інтенціональність) як організуючий процес, що скерований на ціль, можна прирівняти до опису Самості як динамічного патерну, що розгортається в напрямку цілі. Свідомість – це процес, завдяки якому потік динамічних змін, що пронизує Самість, супроводжується якісними відчутними переживаннями.
Емпіричні дані про людину вказують на те, що «мінімальна Самість» пов’язана зі створенням психічних образів та опрацюванням репрезентацій (уявлень), які виринають в результаті «активності в стані спокою» КСС (кіркових внутрішніх(серединних) структур) (Qin and Northoff, 2011; Musholt, 2013; Northoff et al., 2014; Hu et al., 2016; Weiler et al., 2016; LeDoux and Brown, 2017). Більше того, КСС анатомічно та функціонально тісно пов’язані з набором ПКСС (підкорковими структурами середнього мозку) (Northoff and Panksepp, 2008; Panksepp and Northoff, 2009), які вважаються нейрофізіологічним субстратом форми, первісної форми суб’єктивності, названі «Ядерною Самістью» членами цієї дослідницької групи (Panksepp, 1998b) і «прото-Самість» Damasio (1999). Цікаво, що ПКСС є найемоційнішими частинами нашого мозку (можливо, навіть більше, ніж переоцінена лімбічна система), а афективна нейродинаміка, що виникає в ПКСС, відіграє центральну роль в організації людської особистості (Panksepp, 1998b, 2007; Davis et al., 2003; Northoff et al., 2006; Davis and Panksepp, 2011; Panksepp and Davis, 2014; Montag et al., 2016). Отже, як пряме вираження внутрішніх форм функціонування – «намірів у дії» (Panksepp, 1998b) – афекти можуть розглядатися як базові організатори Самості(Panksepp, 1998a,b, 2005, 2010, 2011; Damasio, 1999; Denton, 2006), та виражають форму рудиментарної свідомості (або протосвідомості), яка характеризується первинними процесами та позаоб’єктними (аноетичними) почуттями (Alcaro and Panksepp, 2014).
На початку цієї статті ми цитували текст Юнга, який містить три більш-менш прямо виражені основні тези:
(1) Серед усіх інших юнгіанських архетипів1 є один (Самість), який має центральне значення та універсально розподілений. Він представлений у високоорганізованих геометричних (і символічних) «мандалічних» візерунках (Jung, 1933/1950, 1951) (Малюнок 1).

(2) Якщо існує якась відповідність між мозком і психікою, то фізіологічні основи Самості можуть бути локалізовані в стовбурі мозку, який є основоположною організуючою системою всього мозку та психіки.
(3) Об’єднуючі властивості Самості переважно афективні, окрім ролі впорядкування й орієнтування.
Відповідно, ми думаємо, що такі твердження Юнга не лише були досить далекоглядними, вони насправді відкривають шляхи взваємозв’язку його теорії психики з новітніми науковими теоріями та відкриттями сучасності.
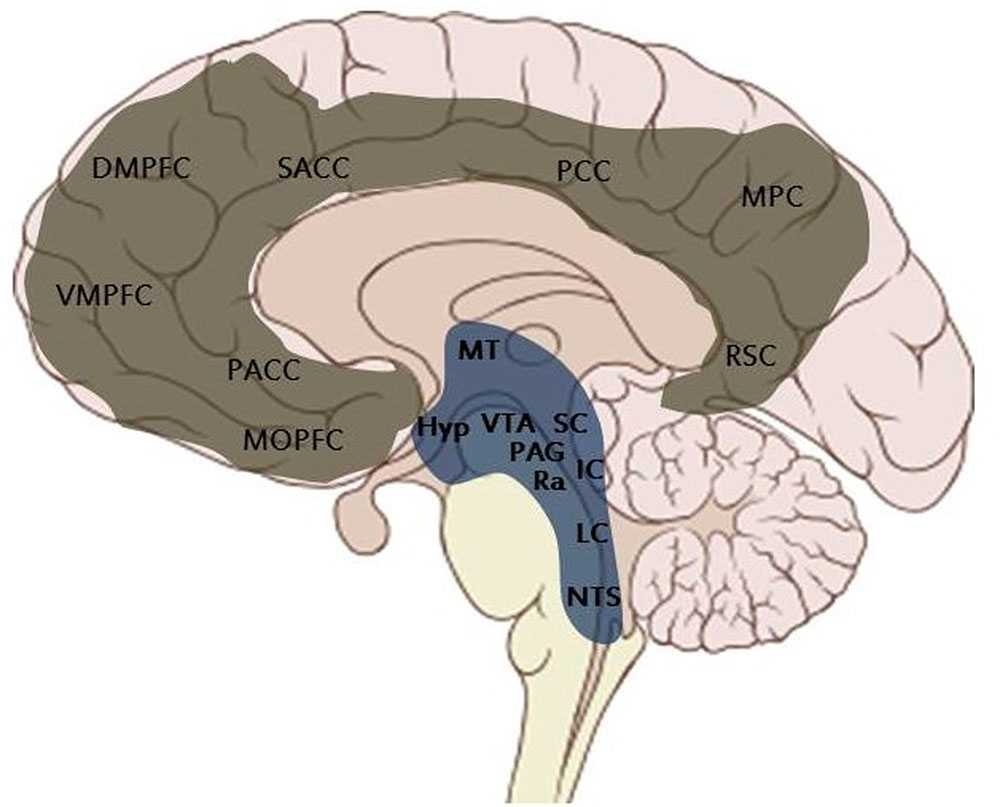
Півкулі головного мозку людини пов’язані зі спинним мозком і рештою тіла через набір ПКСС. Функціональна анатомія ПКСС наближається до зрілості при народженні (Малюнок 2). У своїх новаторських нейроеволюційних роботах MacLean (1990)визначає ПКСС як «рептильний мозок», оскільки вони присутні вже у рептилій і зберігаються досить гомологічно (а не абсолютно незмінно) формі нашарувань від птахів до ссавців. Незважаючи на те, що ця метафора отримала багато критики, ми передбачаємо, що вона означає гомологічно розповсюджений, (але з варіаціями, типовими для різних різновидів) для всіх хребетних «первинний процес).
Дослідження на людях і тваринах показують, що ураження ПКСС викликають кому мозку та припинення будь-якої форми психічного й наміреного життя: вся психічна діяльність руйнується, а організми стають зомбі-подібними, демонструючи переважно вегетативне існування без наміреності (Panksepp, 1998a,b; Watt and Pincus, 2004; Merker, 2007; Panksepp and Biven, 2012). Більше того, дефіцит стає тим важчим, чим нижче по нейро-осі відбувається ушкодження. Наприклад, експериментальні дослідження на тваринах вказують на те, що повне руйнування навколопровідної сірої речовини (PAG), що лежить в основі ПКСС, призводить до повного руйнування опрацювання подій навколишнього середовища, що стосуються саме себе. При повному ушкодженні PAG порушується вся діяльність, спрямована на світ. Тварини перебувають у стані часткової бадьорості, але, здається, вони не можуть виразно усвідомлювати речі (Panksepp and Biven, 2012, стор. 409).
З іншого боку значні ураження кори півкуль та верхніх лімбічних (обідкових) часток не руйнують поле свідомості. Наприклад, тварини, яких позбавили кори на ранніх етапах життя, підтримували чудовий рівень поведінкової узгодженості, наміреності та спонтанності. Вони не лише демонструють здатність вчитися на основі подій із позитивним чи негативним підкріпленням, але й проявляють фактично більшу емоційність, ніж тварини з неушкодженим мозком (Huston and Borbely, 1973, 1974).
Відповідну доказову базу щодо людей сформовано на основі стану, що називається гідроцефалією, при якому кора головного мозку та вищі лімбічні ділянки повністю руйнуються внутрішньоутробно, при цьому більшість підкіркових мереж залишаються функціонуючими. Дивно, але такі діти з гідроцефалією виявляють багато ознак позитивних і негативних афективних станів. Наприклад, вони:
«[…] виражають задоволення посмішкою та сміхом, а відразу – «метушливим» вигином спини та плачем (у багатьох градаціях на їх обличчях зображаються ці емоційні стани). Знайомий дорослий може використовувати таку чуйність у вибудовуванні ігрових послідовностей, які передбачувано прогресують від усмішки, до гиготання, сміху та сильного збудження дитини» (Merker, 2007, стор.79).
Більше того, вони також демонструють певні форми афективного навчання та пам’яті. Вони:
«[…] реалізують поведінкові ініціативи в межах жорстких обмежень їхніх моторної неповносправності у формі інструментальної поведінки, наприклад, створюють галас, б’ючи ногами брязкальця, що висять у спеціальній рамці, сконструйованій для цієї мети («маленька кімната»), або активують улюблені іграшки клавішами – ймовірно, на основі асоціативного засвоєння зв’язку між діями та їхніми наслідками. Така поведінка супроводжується відповідними ситуації ознаками задоволення та збудження з боку дитини» (Merker, 2007).
У підсумку, дані досліджень проведених на тваринах і людях демонструють, що первинна форма Самості виникає в межах ПКСС, і що всі інші складні форми психічного життя можуть залежати від цих областей мозку, оскільки ураження або порушення ПКСС спричиняють колапс усієї свідомої та/або наміреної активності. Незважаючи на той факт, що член цієї дослідницької групи назвали цю структуру «ядерна Самість» (Panksepp, 1998b), а Damasio (1999) назвав її «прото-Самістю», тут ми хотіли б прийняти визначення «Афективне ядро Самості», щоб підкреслити абсолютну актуальність афективного виміру. Дійсно, як наслідок своєї внутрішньої нейрофізіології та нейроанатомічної організації, ПКСС залучені до виникнення основних афективних станів, які диференціюють у межах трьох категорій (Panksepp and Biven, 2012):
(1) Гомеостатичні/вісцеральні афекти відносяться до внутрішніх тілесних станів, які сприймаються на свідомому рівні як загальний настрій і почуття. Дійсно, ПКСС отримують інформацію безпосередньо від організму й, своєю чергою, регулюють вісцеральні та соматичні стани, контролюючи ендокринну, екзокринну й автономну нервову системи 2 (Damasio, 1996, 1999; Porges, 2011). Через жорсткий зв’язок із «внутрішнім середовищем» тіла (Bernard, 1865/1961) ПКСС також називають «вісцеральним мозком» (MacLean, 1990).
(2) Інстинктивні/емоційні афекти відносяться до внутрішніх і найбільш важливих диспозицій що до дій, наближення або уникнення певних ситуацій (див. наступний абзац). Справді, ПКСС еволюційно вже забезпечені нейронними системами, які запускають і контролюють інстинктивні патерни дії та пози, необхідні для виживання та розмноження організму (наприклад, опорно-рухові й орієнтаційні рухи, сексуальну поведінку, поведінку ковтання тощо) (Tinbergen, 1951; Lorenz, 1965; Panksepp, 1998b; Denton, 2006; Arminjon et al., 2010). Деякі з таких інстинктивних патернів складаються з основних емоційних диспозицій, таких як страх, гнів чи радість тощо (Panksepp, 1998b) (див. наступний абзац).
(3) Сенсорні афекти відносяться до рудиментарних перцептивних переживань із внутрішньою афективною цінністю, таких як відчуття дотику, спроможність почути заспокійливі або тривожні звуки, відчуття тепла чи холоду, солодкий чи гіркий смаки тощо. Дійсно, ПКСС забезпечуються деякими первинними сенсорними проміжними станціями для екстероцептивного та пропріоцептивного сприйняття (Merker, 2007). Цікаво, що такі підкоркові зони сприйняття тісно пов’язані з нервовими моторними ядрами, відповідальними за спрямування уваги, такими як верхній горбик та/або оптичний дах, який контролює рухи очей (Stein et al., 2002).
Дослідження мозку з електричною стимуляцією3 дозволили локалізувати в межах ПКСС різноманітні операційні системи мозку ссавців, активація яких відповідає за появу принаймні семи основних емоційних диспозицій (Panksepp, 1998a,b, 2005, 2010; Panksepp and Biven, 2012): системи ПОШУКУ, системи ГНІВУ, системи СТРАХУ, системи ПОХОТІ, системи ПІКЛУВАННЯ, системи ПАНІКИ/СЕПАРАЦІЙНОГО СМУТКУ та системи ГРИ (Малюнок 3). Чотири з таких емоційних систем також були виявлені в мозку рептилій, хоча, можливо, їм бракувало (або вони обмежено розвинені) певних соціальних емоцій, що розвинулися не так давно (ПІКЛУВАННЯ, ПАНІКА/СКОРБОТА та ГРА).

Емоційні операційні системи – це інстинктивні нейронні пристрої, що випускають низку скоординованих та інтегрованих відповідей, які формують характерний репертуар емоцій4. Більше того, дані отримані у дослідженні тварин і людей переконливо свідчать про те, що активація кожної емоційної системи змінює суб’єктивний стан організму та призводить до виникнення характерних почуттів, які сприймаються на свідомому або досвідомому рівні. І справді, люди описують сильні емоційні почуття під час електричної стимуляції ПКСС (Heath, 1964, 1996); тоді як у тварин таке стимулювання завжди викликає ефекти винагороди або відрази, що, ймовірно, впливатиме на їхню поведінку в майбутньому (Panksepp, 2010, 2015).
(До)свідомі характеристики емоцій внутрішньо пов’язані з характером іх наміру, оскільки позитивні та негативні афективні почуття завжди відображають намір наблизитися або уникнути певних ситуацій, що дозволяє визначити емоції як первинні форми диспозицій намірів. Наприклад, диспозиція ПОШУКУ спонукає організм до пошуку основних ресурсів (їжі, води, сексу тощо), СТРАХУ – до уникнення джерела небезпеки тощо5. Отже, емоції завжди виражають диспозиційно-намірену природу, налаштовуючи організм на привілеї світу, таким чином визначаючи основні афективні установки тварин і (в поєднанні з корковими мережами) сфери використання поведінкових стратегій. Така телеологічна точка зору, основоположна й для моделі психіки Юнга. Вона розглядає емоційні афекти як самосприйняття внутрішніх моделей функціонування – «намірів у дії» (Panksepp, 1998b) – що виражає форму «аноетичної» свідомості, що є першим первинним шаром мозку, в якому ядерна Самість афективно переживає власне відчуття себе6.
Відповідно до останніх нелінійних динамічних теорій (Freeman, 1999; Brown, 2002; Llinas, 2002), емоційні афекти можна більш адекватно ідентифікувати як нейродинамічні закономірності, що виникають в межах ПКСС, а потім розповсюджуються по всьому мозку та тілу, зваблюючи цим організм до активності та підштовхуючи його до конкретних «джерел» нейрональної активації (див. також Panksepp, 2000; Krieger, 2014). Таким чином, емоційні афекти діють як вектори, що орієнтують ансамбль поведінкової та психічної активності в напрямку до орбіти значень. Коли вони передаються в напрямку до спинного мозку та інших рухових або вісцеральних ефекторних систем, вони стають інтегрованими ансамблями фізіологічних і поведінкових інстинктивних реакцій. Розповсюджуючись у вищі ділянки мозку, вони можуть приймати форму ансамблів ментальних репрезентацій (архетипових образів або думок), що тяжіють до характерного афективного ядра (Малюнок 4).
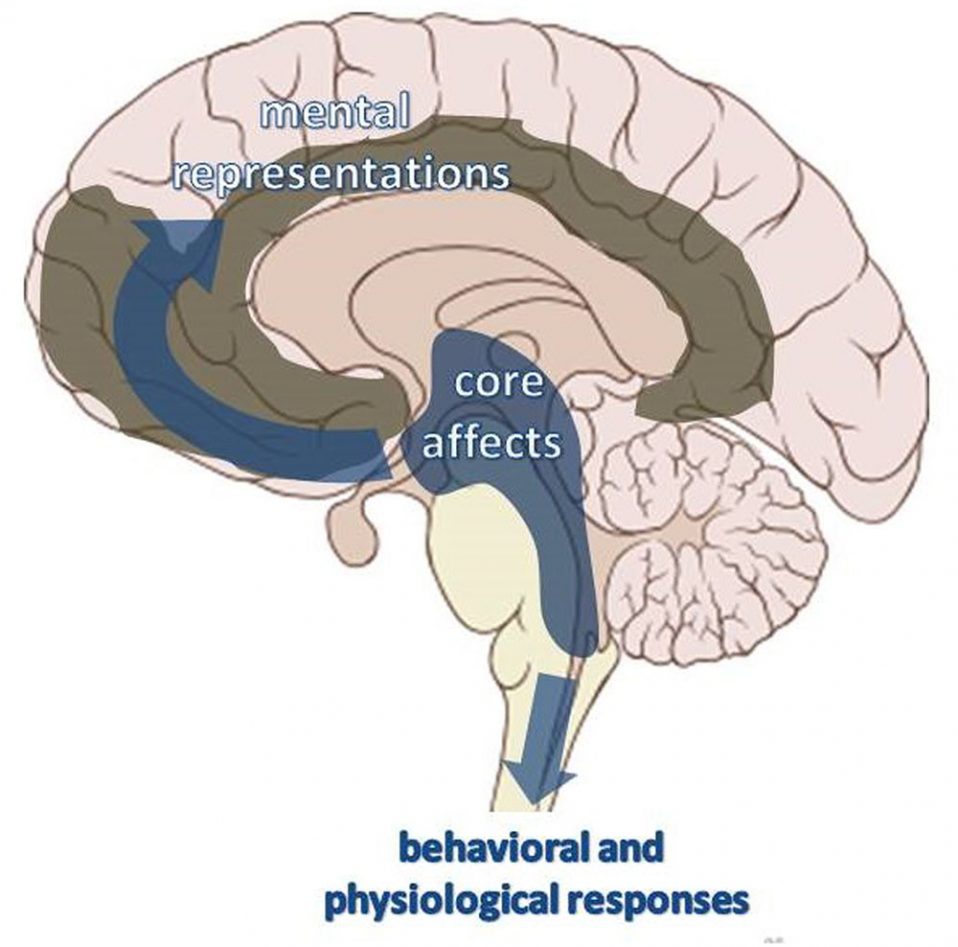
Коли нейродинамічні патерни розвиваються в жорсткий та автоматичний спосіб, вони породжують стереотипні поведінкові або психічні дії, що характеризуються компульсивністю та впливають на суб’єктивний світ так, як зовнішні фактори або, використовуючи юнгіанську термінологію, як «ектопсихічні фактори»:
«Серед психологічних факторів, що визначають поведінку людини, інстинкти є головними силами, що спонукають психічні події. […] Якщо ми розглядатимемо появу психіки як відносно недавню подію в історії еволюції та припустимо, що психічна функція – це явище, що супроводжує нервову систему, яка тим чи іншим чином стала централізованою, то було б важко повірити, що з самого початку інстинкти мали психічний характер. І оскільки зв’язок психіки з мозком є більш вірогідним припущенням, ніж психічна природа життя загалом, я вважаю характеристику компульсивності інстинкту ектопсихічним7 фактором. Тим не менше, він є психологічно важливим, оскільки призводить до формування структур або патернів, які можна розглядати як визначальні фактори поведінки людини» (Jung, 1937/1942, п. 234).
Однак емоційні диспозиції відрізняються від жорстких поведінкових автоматизмів, оскільки ансамбль дій, що координуються кожною емоційною системою, гнучко організується та модулюється. Унаслідок цього емоції осмислюються як «гнучкі патерни дій» (Llinas, 2002), які реагують на тригерні стимули, але також передбачають майбутні події, готують організм до того, щоб справлятися з невизначеними ситуаціями, та, за збереженості неокортексу, орієнтувати його установки відповідно до конкретних намірених шляхів. Внутрішня недетермінована якість емоційних диспозиційних патернів відкриває матеріальну/органічну структуру для входу психічного інтенційного/свідомого зародку (Bergson, 1896/1991). Як підкреслював Юнг:
«[…] Безпосереднім визначальним фактором є не ектопластичний інстинкт, а структура, що виникає в результаті взаємодії інстинкту та поточної психічної ситуації. Таким чином, визначальним фактором буде модифікований інстинкт. Зміни, що їх зазнає інстинкт, настільки ж значущі, як і різниця між кольором, який ми бачимо, й об’єктивною довжиною хвилі, що його продукує. Інстинкт як ектопсихічний фактор просто відіграватиме роль стимулу, тоді як інстинкт як психічний феномен представляв би асиміляцію цього стимулу до вже існуючого патерну. Цей процес слід назвати. Я б назвав його терміном «психізація»8» (Jung, 1937/1942, п. 234).
На поточний історичний момент наукові дослідження не можуть пояснити, як нейродинамічна діяльність ПКСС пов’язана з появою основних афективних станів, які переживаються на суб’єктивному рівні (свідомість) і які можуть телеологічно впливати на хід матеріальних подій (наміреність) (Jonas, 1976; Searle, 1992). Ця складність, ймовірно, виникає, коли суб’єктивні стани (переживання від першої особи) редукціоністично зводять до об’єктивних матеріальних процесів (подій від третьої особи) вдаючись до помилкової спроби помістити інтенсивне та нелокалізоване явище в межах тривимірного просторового поля (Bergson, 1896/1991).
Дуже впливова емпірично орієнтована філософська традиція розглядає афективне почуття як внутрішньо пов’язане зі сприйняттям внутрішніх вісцеральних та тілесних станів явище, своєрідну інтегровану ентероцепцію. Найвідоміша теорія в цьому підході була висунута незалежно James (1884) і Lange (1887). Вони припустили, що афективні почуття відображають кортикально-когнітивну репрезентацію периферично-несвідомого збудження, що активується в організмі певними інстинктивними/емоційними пристроями. Нещодавно Антоніо Дамасіо запропонував нову версію теорії Джеймса-Ланге, виправивши її початковий кортикоцентризм і припустивши, що ПКСС залучені до першого етапу репрезентації соматичних і вісцеральних станів (Damasio, 1996, 1999, 2010).
Концепція Дамасіо має яскравих попередників в історії психоаналізу. Насправді, розглядаючи афекти як стани вісцеральної напруги та розслабленості, Фройд писав:
«Ід, відрізане від зовнішнього світу, має власний світ сприйняття. Воно виявляє з надзвичайною гостротою певні зміни всередині, особливо коливання напруженості своїх інстинктивних потреб, і ці зміни стають усвідомленими як почуття в серіях задоволення-незадоволення» (Freud, 1940/1964, стор. 190).
Нещодавно фройдистські вісцеральні погляди знову підхопили нейропсихоаналітики, які постулюють сувору відповідність між внутрішнім фізіологічним середовищем і психічним світом осіб (Solms and Turnbull, 2002). Згідно з такою точкою зору, афективні почуття пов’язані з процесом гомеостазу (Cannon, 1929; Denton, 2006), а потім із повідомленнями, які тіло посилає мозку-розуму про виживання та репродуктивний успіх чи невдачу. Більше того, останні розробки в нейропсихоаналітичному гомеостатичному підході також свідчать про те, що втілена взаємодія з іншими людьми в дитинстві дозволяє «менталізувати» основні вісцеральні відчуття, перетворюючи грубе внутрішнє сприйняття на суб’єктивні почуття (Fotopoulou and Tsakiris, 2017).
Однак фізіологічний зв’язок ПКСС із внутрішнім і зовнішнім середовищем організму не означає, що афективні почуття просто походять від нейрокогнітивних репрезентацій вісцеральних і соматичних функцій: наприклад, розмірковуючи про природу афектів, сам Фройд зауважував, що скептично ставиться до того, що:
«Цей перелік дозволив нам дійти до суті афекту. Здається, у випадку деяких афектів ми дивимось глибше і усвідомлюємо, що основним […] є повторення певного особливого значущого досвіду. Цей досвід міг бути лише дуже раннім враженням дуже загального характеру, розміщеним у передісторії не окремої людини, а виду» (Фройд, [28], стор. 395).
У цьому надзвичайно значущому фрагменті Фройд відмовляється від своєї традиційної редукційної точки зору, прийнявши ламарківський та антиредукціоністський підхід9. За його словами, внутрішнє ядро афекту – це не просто соматичний матеріальний процес, а психічна подія («дуже раннє враження дуже загального характеру, розміщене в передісторії не окремої людини, а виду»). Тому редукціоністська позиція не повинна виключати ширшої, нередукційної парадигми, яку Юнг постійно розробляв у своїх працях. Мабуть найкращий приклад такого подвійного бачення (редукційного та нередукційного) можна знайти в нарисі «Про природу психічного» (Jung, 1947/1954), де він бере до уваги як біологічні компоненти (наприклад, інстинктивні «патерни поведінки» в тварин і людини), так і їхній зв’язок із непричинними принципами, пов’язаними з фізикою елементарних часток.
Це питання безпосередньо підводить нас до філософської позиції, відомої як «двоаспектний монізм», яку прийняв спочатку Спіноза (див. Ravven, 2013), потім, багато років тому Юнг і Паулі (Atmanspacher, 2012), а нещодавно повторно запропонував Марк Солмс та інші нейропсихоаналітики (Kaplan-Solms and Solms, 2000). Відповідно до такої точки зору, матеріальний і суб’єктивний світи є двома взаємодоповнюючими проявами унікальної, хоча, можливо, й непізнаваної унітарної реальності, яку Юнг описує поняттям «психоїдне». Наявність такого основоположного виміру широко підкреслюється в східній культурній традиції, а також у деяких західних філософів, таких як А.Н. Withehead (1929). У ХХ столітті існування цього виміру було виявлене в ході вивчення квантомеханічних процесів у фізиці та несвідомих процесів в аналітичній психології10. Обидві дисципліни визнавали вплив невидимих (парадоксальних) явищ у межах звичайного потоку видимих матеріальних і психічних подій.
Сьогодні застосування нелінійних динамічних теорій у нейронауці (Freeman, 1999; Brown, 2002; Llinas, 2002; Krieger, 2014), теоретично відкриває шлях до переходу від фізики масивних тіл до квантової фізики, а, отже, до рівня реальності в якій синхроністичні явища можуть бути раціонально прийнятні – зміщуючи рівень неврологічного аналізу з матеріально-нейрохімічного рівня на рівень нематеріально-електричного поля, 11 (Jung, 1955; Bohm, 1980/1981; Penrose, 1989; Brown, 2002). Наприклад, невропатолог Brown (2002), який інтегрував процесуальну філософію Вайтхеда до найновіших відкриттів нейронауки, розглядає Самість як мікрогентичний процес квантових коливань, що піднімається з глибини (підкірки) мозку на поверхню (кортикальний рівень), породжуючи моменти свідомості , які поступово розвиваються від чистого безоб’єктного збудження до складних репрезентацій і саморепрезентації12.
Відповідно до таких спекулятивних гіпотез, ми можемо представити ядерні афекти як «архетипи як такі», що виступають як первинні організовані конфігурації внутрішніх оцінкових подій, які проявляються як у мозково-поведінкових діях/диспозиційних патернах (об’єктивна сфера), так і в інтенсивних афективних почуттях (суб’єктивна сфера). Такі архетипні психоїдні події, ймовірно, в широкому сенсі на фізичному описовому рівні, узгоджуються з динамічною структурою квантових мікрохвиль, що відбуваються в ПКСС13 (див. Brown, 2002). Як ампліфікацію квантових мікрохвиль (Penrose, 1989; Brown, 2002) мозок можна точніше розглядати як трансформаторну станцію, що з’єднує розум і матерію й актуалізує психологічні процеси в лінійному просторово-часовому вимірі. Як пропонував Юнг:
«Можна припустити, що психіка поступово піднімається від низької протяжності до нескінченної напруженності, виходячи за межі, наприклад, швидкості світла, й тим самим дереалізуючи тіло. […] У світлі цього підходу мозок може бути трансформаторною станцією, де відносно нескінченна напруга або напруженість психіки перетворюється на відчутні частоти або «протяжності». І навпаки, згасання інтроспективного сприйняття тіла пояснюється поступовою «психізацією», тобто інтенсифікацією за рахунок розширення. Психіка = найвища інтенсивність у найменшому просторі. НЕПРОТЯЖНА НАПРУЖЕНІСТЬ14»
(C.G. Jung ∼Carl Jung, Letters Vol. II, PP 43–47).
Останні данні нейронауки підтверджують, що афективність є зародковою формою суб’єктивності. Точніше було визнано, що афекти виражають розширену, розсіяну та пов’язану з первинним процесом чутливість, позбавлену будь-якого конкретного змісту або чіткого когнітивного розрізнення між зовнішньо-об’єктивним і внутрішньо-суб’єктивним світом. Таку безоб’єктну та пов’язану з первинним процесом чутливість також називають «аноетичною» свідомістю (або протосвідомістю), «[…] немисленнєвою формою досвіду, яка може бути афективно напруженою, не будучи відомою» (Solms and Panksepp, 2012, стор. 149)15. Це «рудиментарний стан автономної свідомості […] з основоположною формою «самопереживання» від першої особи, що спирається на афективні досвідні стани та сире сенсорне та перцептивне психічне існування» (Vandekerckhove and Panksepp, 2009, стор. 1). У цьому контексті аноетичні протосвідомі почуття виникають як нейродинамічниа «верхівка хвилі» з безперервного потоку дифузного збудження без фокусу чи наміру (настрій) 16.
По суті афективність вважали основоположною рисою мозку-розуму багато ранніх піонерів, включаючи Джона Г’юлінгса Джексона, а також Ойгена Блейлера, одного з найважливіших партнерів у науковій кар’єрі Юнга. Ось що писав Блейлер у своїй знаковій праці про психози:
«[…] У маленької дитини афективність повністю розвинена; всі афекти, які наявні у дорослого, в неї повністю розвинені. І навпаки, інтелект дитини позбавлений змісту, а логічні процеси відносно слабкі» (Bleuler, 1906/1912, стор. 24, переклад наш).
Юнг також, із самого початку (тобто від його «Досліджень асоціацій» (Jung, 1904/1905)) вважав афекти і афективні почуття абсолютно відповідними (релевантними) до суб’єктивного психічного життя, що підкреслюється в наступному уривку:
«Кожний психічний процес має свій, зв’язаний з ним тон почуттів (афективну забарвленість). Це вказує на ступінь впливу процесу на суб’єкта або на те, наскільки цей процес є значущим для нього (в міру того, наскільки цей процес взагалі досягає свідомості). Саме через «афект» суб’єкт стає залученим і відчуває всю вагу реальності» (Jung, 1959/1978, п. 61).
Таким чином Юнг розглядав афекти як сили, які приваблюють суб’єкта та переживаються в формі характерних почуттів. У цьому сенсі Jung (1928/1948) замінив теорію імпульсів (потягів) Фройда набагато переконливішою теорією зв’язуючої сили поля (у психологічному плані – афекту), яка поляризує патерни чи психічні репрезентації та наділяє їх цінністю.
Отже афекти можуть розглядатися як трансперсональні форми досвіду, що пронизуютьь первинне суб’єктивне поле, яке ще не є індивідуальною свідомістью (або самоусвідомленням), оскільки вона живе в недиференційованому континуумі організм/середовище. У той же час афекти є мостом між колективною та індивідуальною психикою, між інстинктивним спадком виду й особистим досвідом, накопиченим в індивідуальному житті. Як описує член цієї дослідницької групи:
«Забезпечуючи спільну нейронну платформу для різноманітного афективного досвіду, ядерна Самість може вважатися номотетичною (універсальною) функцією мозку. Оскільки ядерна Самість поряд з багатьма сирими почуттями, які вона розвиває, взаємодіє з вищими когнітивними […] процесами, вона (Самість) сприяє появі різних ідеографічних (індивідуально унікальних, вдосконалених досвідом) розширених самостей у процесі дозрівання мозку» (Panksepp and Biven, 2012, стор. 392).
Хоча спочатку афекти є безоб’єктними переживаннями, вони забезпечують поле, завдяки якому всі вхідні перцептивні стимули/переживання інтегруються в єдиний свідомий та намірений стан. Більше того, афективні стани здійснюють потужну модуляцію будь-якої форми індивідуального навчання й асоціативної пам’яті (властивість модуляції навчання) (Panksepp and Biven, 2012; Vandekerckhove et al., 2014). Таким чином, весь особистий досвід, накопичений протягом індивідуальної історії, приймає форму кластерів (або комплексів) перцептиних слідів пам’яті (енграм), що тяжіють до афекту. Саме таку ідею розвинув Jung (1960) у вигляді концепції «почуттєво забарвлених комплексів», які він розглядав як психологічні структури, що об’єднують різні психічні змісти та репрезентації на основі спільного афективного стану17. Кожен комплекс об’єднаний тією самою емоцією, яка визначає ядро його змісту й організовує досвід, сприйняття й афект навколо постійної центральної теми. Наприклад, комплекс меншовартості – це констеляція спогадів, думок і фантазій, пов’язаних із браком самоцінності, сумнівами та невпевненістю в собі, а також почуттями невідповідності стандартам.
Так, говорячи про комплекс, Юнг писав: «Констелююча сила його ядерного елемента відповідає його ціннісній напруженості, тобто його енергії» (Jung, 1928/1948, §19, курсив Юнга). У цьому ж дусі він також писав: «Це було б непростимим гріхом та спрощенням, якби ми не помічали почуттєвої цінності архетипу. Це надзвичайно важливо як теоретично, так і терапевтично» [ЗТ 8, п. 411], оскільки архетип – це «[…] жива система реакцій і схильностей», пов’язана з живою особистістю «[…] містком емоцій» [ЗТ 18, п. 589].
Юнгіанська теорія почуттєво забарвлених комплексів є розробкою роботи П’єра Жане щодо автономних фіксованих ідей. Відповідно до Janet (1889), фіксовані ідеї – це ментальні образи або думки, що мають високий емоційний заряд і набувають перебільшених пропорцій, тому вони можуть не бути інтегрованими до его-свідомості звичним чином, та ставати ізольованими від звичної особистості, створюючи дисоційовані стани розуму (Monahan, 2009). Однак, на відміну від оригінальної теорії Жане, Юнг стверджував, що дисоціативний аспект комплексів, як правило, зворотний, тому вони можуть в тій чи іншій мірі інтегруватися відповідно до поточної ситуації. Тільки за важких психічних патологій, таких як психози, певні комплекси назавжди дисоціюються від свідомого «Я», й особистість стає фрагментованою.
Багато авторів висували гіпотезу, що Самість не є цілісним явищем, і що її можна диференціювати як нашарування, які формувалися в процесі еволюції (James, 1890/1950). Психоаналітики широко використовували археологічну метафору для опису людського розуму. Наприклад, Зигмунд Фройд бачив психіку як древнє місто, як-от Рим, де хід часу відбився в монументальних стратифікаціях від античності до сучасної епохи. Дослідження багатошарової організації людської психики були емпірично збагачені завдяки внеску нейронаук. Джон Гаглінгс Джексон, батько англійської неврології, був, мабуть, першим нейрофізіологом, який запропонував чіткий ієрархічний опис функціонування мозку (див. Franz and Gillett, 2011), а його цікаві здогадки отримали подальший розвиток у роботах Lurija (1962). Більше того, інтегруючи нейроанатомічні погляди на людину та тварин, MacLean (1990) запропонував важливий нейроеволюційний погляд на сучасну нейронауку (хоча й не без широких дискусій – див. їх огляд, Panksepp, 2002). Він концептуалізував три шари, що взаємно накладаються в мозку людини: неокортикально-раціональний мозок, характерний для нашого виду, проміжний лімбічно-емоційний мозок, характерний для всіх ссавців, і вісцерально-інстинктивний мозок, характерний для рептилій – патерн, який більш-менш зберігся серед хребетних18.
Утім, на відміну від домінуючої антропоцентричної точки зору, доказова база нейронауки, що накопичується, підтверджує, що первинна форма суб’єктивності вже присутня на першому рівні ієрархії мозку (інстинктивний мозок рептилій-древніх ссавців), і що вона широко розповсюджена серед ссавців, птахів і, можливо, інших хребетних (Panksepp, 1998b, 2016; Seth et al., 2005; Northoff and Panksepp, 2008; Edelman and Seth, 2009; Panksepp and Northoff, 2009; Revonsuo, 2010; Ward, 2011; Panksepp and Biven, 2012; Fabbro et al., 2015; Feinberg and Mallatt, 2016). Зростаючі докази змусили члена цієї дослідницької групи запропонувати певні кардинальні зміни до трискладової нейроеволюційної моделі Пола Макліна19 (Panksepp, 1998b, 2011; Panksepp and Biven, 2012) та спонукали сучасний нейропсихоаналітичний рух розглядати реальність афективного (якщо не когнітивно-рефлексивного) «свідомого ІД»20 (Solms and Panksepp, 2012; Solms, 2013).
Відповідно до таких гіпотез нещодавно було запропоновано думку, що явище свідомості можна диференціювати як аноетичні, ноетичні та автоноетичні стани, що відповідають різним шарам нервово-психічної архітектури (Vandekerckhove and Panksepp, 2009, 2011).
Аноетична свідомість (вже описана в попередніх абзацах) – це перший первинний шар Самості21, древній нейроеволюційний продукт, широко розповсюджений серед видів тварин, починаючи від ссавців і закінчуючи птахами та плазунами та, можливо, усіма хребетними (Fabbro et al., 2015; Feinberg and Mallatt, 2016). Більше того, новонароджені приходять у світ з успадкованою здатністю переживати аноетичні стани в результаті діяльності підкіркових областей мозку, які вже функціонально дозрілі при народженні (Merker, 2007), як ми описали в цій статті.
З іншого боку, ноетична й автоноетична свідомість є новішими еволюційними функціями вищих кіркових і лімбічних ділянок мозку та потребують для адекватного розвитку та вираження певної форми індивідуального навчання.
Ноетична свідомість відображає досвід, накопичений на основі знань досвід, який виникає, коли вдосконалені здатності уваги дозволяють чітко розмежувати та класифікувати специфічні особливості середовища, що при достатній збереженності неокортексу дозволяє тваринам мислити наперед. Справді, коли в центрі уваги опиняються конкретні аспекти подій, на перший план виходить експліцитне об’єктне рефлексивне усвідомлення, тоді як семантична (концептуальна) пам’ять допомагає аналізувати та класифікувати ситуацію (Tulving, 1985). Ця форма свідомості, яку Едельман назвав «первинною свідомістю», та яку він пов’язав з діяльністю зворотніх таламо-кортикальних мозкових схем (Edelman, 1989). Непрямі дані свідчать про те, що ноетична свідомість присутня в ссавців, птахів і, можливо, також у плазунів (Edelman et al., 2005), і що людські немовлята, народжені дуже незрілими, починають проявляти її після третього місяця розвитку.
Наступний крок у нейро-психічній еволюції здійснюється шляхом засвоєння мови (Hauser et al., 2002; Gazzaniga, 2011) та здатності подумки подорожувати в часі (Corballis, 2012), що робить можливим не лише свідоме згадування минулих подій (епізодичну пам’ять), а й уявлення майбутніх подій, включаючи власну смерть (Tulving, 2002). Ця наративна структура Самості, яка, ймовірно, з’явилася понад 1 мільйон років тому, найбільш чітко (для нас) проявилася в роді Homo habilis, породжує найскладнішу автоноетичну свідомість на основі самосвідомості, яку Едельман описав як «вторинну свідомість» (Edelman, 1992). Цікаво, що автоноетична свідомість, здається, тісно пов’язана з так званою «теорією розуму» (здатністю уявляти, що відбувається в свідомості інших людей), яка, очевидно, є в основному рисою людини (Corballis, 2012).
Перехід від афективної ядерної Самості (аноетичної свідомості) до вищих стадій еволюції Самості (ноетична й автоноетична свідомість) – це еволюційний стрибок, який здійснили люди, стрибок, який давно був підготовлений попередніми еволюційними процесами в інших ссавців і, можливо, птахів та інших істот. У наступних роботах ми дослідимо й опишемо такі еволюційні досягнення, а також новий творчий потенціал, що його відкриває набуття когнітивно-рефлексивних спроможностей22. Тут ми зосередилися на найосновніших засадах нашого буття – афективних субстратах, з яких походять усі наші вищі психологічні функції. Ці початкові шари нашого розуму забезпечують важливу основу для всіх наступних психоневрологічних функцій вищого порядку, які роблять людський розум унікальним.
Хоча домінантні нейрокогнітивні парадигми, як правило, співвідносять суб’єктивне життя з найвищими рівнями мозкової організації (головним чином із накопиченням індивідувальних спогадів, що зберігаються в нейропластичних ланцюгах переднього мозку), великий обсяг нейроетологічних доказів показує, що тварини, які не є людьми (ссавці, птахи та, можливо, також інші хребетні) також мають форми суб’єктивності, що виникають внаслідок діяльності давнього еволюційного підкіркового стовбуру мозку, проміжного мозку та базальної ділянки переднього мозку (Panksepp, 1998b, 2015, 2016; Seth et al., 2005; Edelman and Seth, 2009; Revonsuo, 2010; Ward, 2011; Panksepp and Biven, 2012; Fabbro et al., 2015). Ці висновки чітко вказують на те, що суб’єктивність – це успадкована диспозиція, спрямований на інстинктивні архаїчні основи дії нашого мозку (Goodwyn, 2010), й вони підтверджують думку Юнга про те, що до того, як в процесі розвитку немовлята набувають рефлексивну самосвідомість, вже існує первісно-інстинктивна афективна форма Самості, що виражається у формі афективно-психічної інтенційності, яка може ефективно, оціночно взаємодіяти з матеріальним, детермінованим світом.
У своїй відомій автобіографії (Jung, 1962/1963) Карл Г. Юнг описує власний сон, який він вважав найважливішим одкровенням щодо структури психіки:
«Я був у незнайомому мені будинку, в якому було два поверхи. Це був «мій дім». Я опинився на верхньому поверсі, де була ніби вітальня, омебльована вишуканими старовинними предметами в стилі рококо. На стінах висіли безліч дорогоцінних старих картин. Я дивувався, що це мій будинок, і думав: «Непогано». Але потім мені спало на думку, що я не знаю, як виглядає нижній поверх.
Спустившись сходами, я дійшов до першого поверху. Там усе було набагато старіше. Я усвідомив, що ця частина будинку має датуватися приблизно XV або XVI століттям. Меблі були середньовічні, підлоги – з червоної цегли. Скрізь було досить темно. Я переходив з однієї кімнати до іншої, думаючи: «Тепер я дійсно повинен дослідити весь будинок». Я натрапив на важкі двері й відчинив їх. За ними я виявив кам’яні сходи, які вели вели до льоху.
Знову спустившись, я опинився в красивому склепінчастому приміщенні, яке виглядало надзвичайно старовинним. Оглядаючи стіни, я побачив шари цегли між звичайних кам’яних блоків й уламки цегли в вапняному розчині. Щойно я це побачив, я зрозумів, що ці стіни походять із римських часів. Тепер мій інтерес став дуже сильним. Я пильніше придивився до підлоги. Це були кам’яні брили, й в одній із них я виявив кільце. Коли я потягнув його, кам’яна плита піднялася, і я знову побачив вузькі кам’яні сходи, що вели вниз у глибину.
Ними я теж спустився й увійшов до низької печери, вирізаної в скелі. На підлозі лежав густий пил, а в пилу були розкидані кістки та зламана кераміка, ніби залишки первісної культури. Я виявив два людські черепи – очевидно, дуже старі, вони напіврозпалися. Потім я прокинувся» (Jung, 1962/1963, стор. 155).
Таке сновидіння – ще один приклад того, як психіка може складатися з безлічі нервових шарів, коли поверхневі та недавні шари формуються на основі старших шарів – сформованих протягом розвитку природної (та культурної) історії нашого виду. Розмірковуючи про значення найглибшого та зачаткового шару, печери, Юнг писав:
«[Тут] ми сягаємо оголеної кам’яної породи, а разом із нею й того доісторичного часу, коли мисливці на північних оленів боролися за своє голе та жалюгідне існування зі стихіями дикої природи. Людьми тієї епохи все ще повністю володіли тваринні інстинкти, без яких життя було б неможливим» (63, п. 55).
У Антоніо Алькаро з’явилася оригінальна ідея, він написав перший проект рукопису. Стефано Карта зробив важливий теоретичний і клінічний внесок, особливо стосовно юнгіанського підходу. Яак Панксепп працював над основними нейронауковими частинами та редагував весь рукопис.
Автори заявляють, що дослідження проводилось за відсутності будь-яких комерційних або фінансових відносин, які можна було б трактувати як потенційний конфлікт інтересів.
Після болісної смерті Яака Панксеппа (18 квітня 2017 р.) Антоніо Алькаро та Стефано Карта хочуть присвятити цю статтю пам’яті друга та безцінного колеги.
КСС – кортикальні серединні структури; ПКСС – підкіркові серединні структури.
Alcaro, A., and Panksepp, J. (2014). “Le radici affettive e immaginative del Sé. Un’indagine neuroetologica sulle origini della soggettività,” in La Plasticità del Sé. Una Prospettiva Neuropsicodinamica, eds G. Nortoff, M. Farinelli, R. Chattat, and F. Baldoni (Bologna: Il Mulino), 65–89.
Arminjon, M., Ansermet, F., and Magistretti, P. (2010). The homeostatic psyche: freudian theory and somatic markers. J. Physiol. Paris 104, 272–278. doi: 10.1016/j.jphysparis.2010.08.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Atmanspacher, H. (2012). Dual-aspect monism à la pauli and jung. J. Conscious. Stud. 19, 96–120.
Bergson, H. (1896/1991). Matter and Memory. New York, NY: Zone Books.
Bernard, C. (1865/1961). Introduction to the Study of Experimental Medicine. New York, NY: Collier.
Bleuler, E. (1906/1912). Affectivity, Suggestibility, Paranoia, Trans. C. Ricksher. Utica, NY: State Hospitals Press.
Bohm, D. (1980/1981). Wholeness and the Implicate Order. Abingdon: Routledge & Kegan Paul.
Bollas, C. (1987). The Shadow of the Object: Psychoanalysis of the Unthought Known. New York, NY: Columbia University Press.
Brown, J. W. (2002). The Self-Embodying Mind. Process, Brain Dynamics and the Conscious Present. New York, NY: Barrytown.
Cannon, W. B. (1929). Organization for physiological homeostasis. Physiol. Rev. 9, 399–431.
Carhart-Harris, R., and Friston, K. (2010). The default-mode, ego-functions and free-energy: a neurobiological account of Freudian ideas. Brain 133, 1265–1283. doi: 10.1093/brain/awq010
PubMed Abstract | CrossRef Full Text | Google Scholar
Corballis, M. C. (2012). The wandering mind: mental time travel, theory of mind and language. Anál. Soc. 205, 870–893. doi: 10.1511/2012.96.210
CrossRef Full Text | Google Scholar
Crick, F. (1994). The Astonishing Hypothesis: The Scientific Search for the Soul. New York, NY: Charles Scribner’s Sons.
Damasio, A. (1999). The Feeling of What Happens: Body and Emotion in the Making of Consciousness. New York, NY: Harcourt Brace.
Damasio, A. (2010). The Self Comes Into Mind. Constructing the Conscious Brain. New York, NY: Pantheon, Hardcover.
Damasio, A. R. (1996). The somatic marker hypothesis and the possible functions of the prefrontal cortex. Philos. Trans. R. Soc. Lond. B Biol. Sci. 351, 1413–1420. doi: 10.1098/rstb.1996.0125
PubMed Abstract | CrossRef Full Text | Google Scholar
Davis, K., Panksepp, J., and Normansell, L. (2003). The affective neuroscience personality scales. J. Neuropsychoanal. 5, 57–70. doi: 10.1080/15294145.2003.10773410
CrossRef Full Text | Google Scholar
Davis, K. L., and Panksepp, J. (2011). The brain’s emotional foundations of human personality and the affective neuroscience personality scales. Neurosci. Biobehav. Rev. 35, 1946–1958. doi: 10.1016/j.neubiorev.2011.04.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Denton, D. A. (2006). The Primordial Emotions: The Dawning of Consciousness. New York, NY: Oxford University Press. doi: 10.1093/acprof:oso/9780199203147.001.0001
CrossRef Full Text | Google Scholar
Eccles, J. C. (1994). How the Self Controls its Brain. Berlin: Springer.
Edelman, D. B., Baars, B. J., and Seth, A. K. (2005). Identifying hallmarks of consciousness in non-mammalian species. Conscious. Cogn. 14, 169–187. doi: 10.1016/j.concog.2004.09.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Edelman, D. B., and Seth, A. K. (2009). Animal consciousness: a synthetic approach. Trends Neurosci. 32, 476–484. doi: 10.1016/j.tins.2009.05.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Edelman, G. M. (1989). The Remembered Present: A Biological Theory of Consciousness. New York, NY: Basic Books.
Edelman, G. M. (1992). Bright Air, Brilliant Fire: On the Matter of the Mind. New York, NY: Basic Books.
Edelman, G. M., and Tononi, G. (2000). Consciousness: How Matter Becomes Imagination. London: Allen Lane.
Fabbro, F., Aglioti, S. M., Bergamasco, M., Clarici, A., and Panksepp, J. (2015). Evolutionary aspects of self- and world consciousness in vertebrates. Front. Hum. Neurosci. 9:157. doi: 10.3389/fnhum.2015.00157
PubMed Abstract | CrossRef Full Text | Google Scholar
Feinberg, T. F., and Mallatt, J. M. (2016). The Ancient Origins of Consciousness. Cambridge MA: MIT Press.
Fonagy, P. (2001). Attachment Theory and Psychoanalysis. New York, NY: Other Press.
Fotopoulou, A., and Tsakiris, M. (2017). Mentalizing homeostasis: the social origins of interoceptive inference. Neuropsychoanalysis 19, 3–28. doi: 10.1080/15294145.2017.1294031
CrossRef Full Text | Google Scholar
Franz, E. A., and Gillett, G. (2011). John Hughlings Jackson’s evolutionary neurology: a unifying framework for cognitive neuroscience. Brain 134, 3114–3120. doi: 10.1093/brain/awr218
PubMed Abstract | CrossRef Full Text | Google Scholar
Freeman, W. J. (1999). How Brains Make Up Their Minds. London: Weidenfeld and Nicolson.
Freud, S. (1940/1964). “An outline of psychoanalysis,” in The Standard Edition of the Complete Psychological Works of Sigmund Freud, Vol. 32, ed. J. Strachey (London: Horgart Press).
Gazzaniga, M. S. (2011). Who’s in Charge? Free Will and the Science of the Brain. New York, NY: Ecco.
Goodwyn, E. (2010). Approaching archetypes: reconsidering innateness. J. Anal. Psychol. 55, 502–521. doi: 10.1111/j.1468-5922.2010.01862.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Hauser, M. D., Chomsky, N., and Fitch, W. T. (2002). The faculty of language: what is it, who has it, and how did it evolve? Science 298, 1569–1579.
Heath, R. G. (1964). “Pleasure response of human subjects to direct stimulation of the brain: physiologic and psychodynamic considerations,” in The Role of Pleasure in Human Behavior, ed. R. G. Heath (New York, NY: Hoeber), 219–243.
Heath, R. G. (1996). Exploring the Mind-Body Relationship. Baton Rouge LA: Moran Printing.
Heisenberg, W. (1958/2000). Physics and Philosophy: The Revolution in Modern Science. London: Penguin Books.
Hu, C., Di, X., Eickhoff, S. B., Zhang, M., Peng, K., Guo, H., et al. (2016). Distinct and common aspects of physical and psychological self-representation in the brain: a meta-analysis of self-bias in facial and self-referential judgements. Neurosci. Biobehav. Rev. 61, 197–207. doi: 10.1016/j.neubiorev.2015.12.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Huston, J. P., and Borbely, A. A. (1973). Operant conditioning in forebrain ablated rats by use of rewarding hypothalamic stimulation. Brain Res. 50, 467–472. doi: 10.1016/0006-8993(73)90753-1
CrossRef Full Text | Google Scholar
Huston, J. P., and Borbely, A. A. (1974). The thalamic rat: general behavior, operant learning with rewarding hypothalamic stimulation, and effects of amphetamine. Physiol. Behav. 12, 433–448.
PubMed Abstract | Google Scholar
James, W. (1884). What is an emotion? Mind 9, 188–205. doi: 10.1093/mind/os-IX.34.188
CrossRef Full Text | Google Scholar
James, W. (1890/1950). The Principles of Psychology. New York, NY: Dover.
Janet, P. (1889). L’Automatisme Psychologique. Paris: L’Harmattan.
Jonas, H. (1976). “On the power or impotence of subjectivity,” in Philosophical Dimensions of the Neuro-Medical Sciences, eds S. F. Spicker and H. T. Engelhardt (Dordrecht: Reidel Publishing).
Jouvet, M. (1975). “The function of dreaming. A neurophysiologist’s point of view,” in Handbok of Psychology, eds M. S. Gazzaniga and C. Blackemore (New York, NY: University Press).
Jung, C. G. (ed.) (1951). “The structure and dynamics of the Self,” in The Collected Works, Vol. 9, (Princeton, NJ: Princeton University Press).
Jung, C. G. (ed.) (1955). “Synchronicity: an acausal connecting principle,” in The Collected Works, 2nd Edn, Vol. 8, (Princeton, NJ: Princeton University Press).
Jung, C. G. (ed.) (1958). “The psychogenesis of mental disease,” in The Collected Works, Vol. 3, (Princeton, NJ: Princeton University Press).
Jung, C. G. (ed.) (1960). “The structure and dynamics of the psyche,” in The Collected Works, Vol. 8, (Princeton, NJ: Princeton University Press).
Jung, C. G. (1962/1963). Memories, Dreams, Reflections. New York, NY: Pantheon Books.
Jung, C. G. (ed.) (1904/1905). “Experimental researches,” in The Collected Works, Vol. 2, (Princeton, NJ: Princeton University Press).
Jung, C. G. (ed.) (1928/1948). “The psychological foundations of the belief in spirits,” in The Collected Works, Vol. 8, (Princeton, NJ: Princeton University Press).
Jung, C. G. (ed.) (1933/1950). “A study in the process of individuation,” in The Collected Works, Vol. 9, (Princeton, NJ: Princeton University Press).
Jung, C. G. (ed.) (1937/1942). “Psychological factors in human behavior,” in The Collected Works, Vol. 8, (Princeton, NJ: Princeton University Press).
Jung, C. G. (ed.) (1947/1954). “On the nature of the psyche,” in The Collected Works, Vol. 8, (Princeton, NJ: Princeton University Press).
Jung, C. G. (ed.) (1959/1978). “Aion,” in The Collected works, Vol. 9, (Princeton, NJ: Princeton University Press).
Kandel, E. R. (2005). Psychiatry, Psychoanalysis, and the New Biology of Mind. Washington, DC: American Psychiatric Publishing.
Kaplan-Solms, K., and Solms, M. (2000). Clinical Studies in Neuro-Psychoanalysis: Introduction to a Depth Neuropsychology. London: Karnac.
Kauffman, S. (1993). The Origins of Order. Self Organization and Selection in Evolution. Oxford: Oxford University Press.
Krieger, N. M. (2014). Bridges to Consciousness. Complexes and Complexity. New York, NY: Routledge.
Lange, C. (1887). Ueber Gemüthsbewegungen. Leipzig: Theodor Thomas.
LeDoux, J. E. (1996). The Emotional Brain. New York, NY: Simon and Schuster.
LeDoux, J. E. (2015). Anxious: Using the Brain to Understand and Treat Fear and Anxiety. New York, NY: Viking.
LeDoux, J. E., and Brown, R. (2017). A higher-order theory of emotional consciousness. Proc. Natl. Acad. Sci. U.S.A. 114, E2016–E2025. doi: 10.1073/pnas.1619316114
PubMed Abstract | CrossRef Full Text | Google Scholar
Llinas, R. (2002). I of the Vortex: From Neurons to Self. Cambridge MA: MIT Press.
Lorenz, K. (1965). Evolution and Modification of Behavior. London: Metheuen.
Lurija, A. R. (1962). Higher Cortical Functions in Man. Moscow: Moscow University Press.
MacLean, P. D. (1990). The Triune Brain in Evolution: Role in Paleocerebral Functions. New York, NY: Plenum Press.
McFadden, J., and Al-Khalili, J. (2014). Life on the Edge: The Coming of Age of Quantum Biology. New York City, NY: Random House.
McGinn, C. (1989). Can we solve the mind-body problem? Mind 98, 349–366. doi: 10.1093/mind/XCVIII.391.349
CrossRef Full Text | Google Scholar
Merker, B. (2007). Consciousness without a cerebral cortex: a challenge for neuroscience and medicine. Behav. Brain Sci. 30, 63–81. doi: 10.1017/S0140525X07000891
PubMed Abstract | CrossRef Full Text | Google Scholar
Monahan, P. (2009). C.G.Jung: Freud’s heir or Janet’s? The influence upon Jung of Janet’s dissociationism. Int. J. Jungian Stud. 1, 33–49. doi: 10.1080/19409050802681876
CrossRef Full Text | Google Scholar
Montag, C., Hahn, E., Reuter, M., Spinath, F. M., Davis, K., and Panksepp, J. (2016). The role of nature and nurture for individual differences in primary emotional systems: evidence from a twin study. PLoS ONE 11:e0151405. doi: 10.1371/journal.pone.0151405
PubMed Abstract | CrossRef Full Text | Google Scholar
Moruzzi, G., and Magoun, H. W. (1949). Brain stem reticular formation and activation of the EEG. Electroencephalogr. Clin. Neurophysiol. 1, 455–473.break doi: 10.1016/0013-4694(49)90219-9
CrossRef Full Text | Google Scholar
Musholt, K. (2013). A philosophical perspective on the relation between cortical midline structures and the self. Front. Hum. Neurosci. 7:536. doi: 10.3389/fnhum.2013.00536
PubMed Abstract | CrossRef Full Text | Google Scholar
Northoff, G. (2013). Unlocking the Brain: Consciousness, Vol. 2. Oxford: Oxford University Press.
Northoff, G., Farinelli, M., Chattat, L., and Baldoni, F. (2014). La Plasticità del Sè. Un Approccio Neuropsicodinamico. Bologna: Il Mulino.
Northoff, G., Heinzel, A., de Greck, M., Bermpohl, F., Dobrowolny, H., and Panksepp, J. (2006). Self-referential processing in our brain–a meta-analysis of imaging studies on the self. Neuroimage 31, 440–457. doi: 10.1016/j.neuroimage.2005.12.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Northoff, G., and Panksepp, J. (2008). The trans-species concept of self and the subcortical-cortical midline system. Trends Cogn. Sci. 12, 259–264. doi: 10.1016/j.tics.2008.04.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Panksepp, J. (1998a). The periconscious substrates of consciousness: affective states and the evolutionary origins of the SELF. J. Conscious. Stud. 5, 566–582.
Panksepp, J. (1998b). Affective Neuroscience: The Foundations of Human and Animal Emotions. New York, NY: Oxford University Press.
Panksepp, J. (2000). “The neurodynamics of emotions: an evolutionary-neurodevelopmental view,” in Emotion, Self-Organization, and Development, eds M. D. Lewis and I. Granic (New York, NY: Cambridge University Press), 236–264.
Panksepp, J. (2002). “The MacLean legacy and some modern trends in emotion research,” in The Evolutionary Neuroethology of Paul MacLean, eds G. A. Cory Jr. and R. Gardner Jr. (Westport, CT: Praeger), 9–25.
Panksepp, J. (2005). Affective consciousness: core emotional feelings in animals and humans. Conscious. Cogn. 14, 30–80. doi: 10.1016/j.concog.2004.10.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Panksepp, J. (2007). “Affective consciousness,” in The Blackwell Companion to Consciousness, eds M. Velmans and S. Schneider (Malden, MA: Blackwell Publishing), 114–129.
Panksepp, J. (2010). Affective consciousness in animals: perspectives on dimensional and primary process emotion approaches. Proc. Biol. Sci. 277, 2905–2907. doi: 10.1098/rspb.2010.1017
PubMed Abstract | CrossRef Full Text | Google Scholar
Panksepp, J. (2011). Cross-species affective neuroscience decoding of the primal affective experiences of humans and related animals. PLoS ONE 6:e21236. doi: 10.1371/journal.pone.0021236
PubMed Abstract | CrossRef Full Text | Google Scholar
Panksepp, J. (2015). “The neuroevolutionary sources of mind,” in The Constitution of Phenomenal Consciousness: Toward a Science and Theory, ed. S. M. Miller (Amsterdam: John Benjamins), 226–259.
Panksepp, J. (2016). The cross-mammalian neurophenomenology of primal emotional affects: From animal feelings to human therapeutics. J. Comp. Neurol. 524, 1624–1635. doi: 10.1002/cne.23969
PubMed Abstract | CrossRef Full Text | Google Scholar
Panksepp, J., and Biven, L. (2012). The Archaeology of Mind: Neuroevolutionary Origins of Human Emotion. New York, N Y: W. W. Norton & Company.
Panksepp, J., and Davis, K. (2014). The emotional fundamentals of personality and the higher affective polarities of mind: comment on “personality from a cognitive-biological perspective” by Y. Neuman. Phys. Life Rev. 11, 691–692. doi: 10.1016/j.plrev.2014.10.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Panksepp, J., and Northoff, G. (2009). The trans-species core SELF: the emergence of active cultural and neuro-ecological agents through self-related processing within subcortical-cortical midline networks. Conscious. Cogn. 18, 193–215. doi: 10.1016/j.concog.2008.03.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Penrose, R. (1989). The Emperor’s New Mind. New York, NY: Oxford University Press.
Porges, S. W. (2011). The Polyvagal Theory: Neurophysiological Foundations of Emotions, Attachment, Communication, and Self-regulation. New York, NY: W. W. Norton.
Qin, P., and Northoff, G. (2011). How is our self related to midline regions and the default-mode network? Neuroimage 57, 1221–1233. doi: 10.1016/j.neuroimage.2011.05.028
PubMed Abstract | CrossRef Full Text | Google Scholar
Ravven, H. M. (2013). The Self Beyond Itself: An Alternative History of Ethics, the New Brain Sciences, and the Myth of Free Will. New York, NY: The New Press.
Revonsuo, A. (2010). Consciousness. The Science of Subjectivity. Hove: Psychology Press.
Rolls, E. (1999). The Brain and Emotion. New York, NY: Oxford University Press.
Sander, L. (2007). Sistemi Viventi. Milan: Raffaello Cortina.
Schore, A. N. (1994). Affect Regulation and the Origin of the Self. Mahwah, NJ: Erlbaum.
Schrödinger, E. (1944). What Is Life? the Physical Aspect of the Living Cell – Mind and Matter. Cambridge: Cambridge University Press.
Searle, J. (1992). The Rediscovery of the Mind. Cambridge: MIT Press.
Searle, J. R. (1991). Consciousness, Unconsciousness, and Intentionality. Philos. Issues 1, 45–66. doi: 10.2307/1522923
CrossRef Full Text | Google Scholar
Seth, A., Baars, B. J., and Edelman, D. B. (2005). Criteria for consciousness in humans and other mammals. Conscious.Cogn. 14, 119–139. doi: 10.1016/j.concog.2004.08.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Simondon, G. (1989/2007). L’individuation Psychique et Collective. Paris: Aubier.
Solms, M. (2013). The conscious “Id”. Neuropsychoanalysis 15, 5–20. doi: 10.1080/15294145.2013.10773711
CrossRef Full Text | Google Scholar
Solms, M., and Panksepp, J. (2012). The “id” knows more than the “ego” admits: neuropsychoanalytic and primal consciousness perspectives on the interface between affective and cognitive neuroscience. Brain Sci. 2, 147–175. doi: 10.3390/brainsci2020147
PubMed Abstract | CrossRef Full Text | Google Scholar
Solms, M., and Turnbull, O. (2002). The Brain and the Inner World: An Introduction to the Neuroscience of Subjective Experience. New York, NY: Other Press.
Stein, B. E., Wallace, M. W., Stanford, T. R., and Jiang, W. (2002). Cortex governs multisensory integration in the midbrain. Neuroscientist 8, 306–314. doi: 10.1177/107385840200800406
PubMed Abstract | CrossRef Full Text | Google Scholar
Stern, D. N. (1985). The Interpersonal World of the Infant. New York, NY: Basic Books.
Stevens, A. (2003). The Archetype Revisited. An Updated Natural Hystory of the Self. Toronto: Inner City Book.
Tagliagambe, S., and Malinconico, A. (2011). Pauli e Jung. Un Confrnto su Materia e Psiche. Milan: Raffaello Cortina.
Tinbergen, N. (1951). The Study of Instinct. Oxford: Oxford University Press.
Tulving, E. (1985). Memory and consciousness. Canad. Psychol. 26, 1–12. doi: 10.1037/h0080017
CrossRef Full Text | Google Scholar
Tulving, E. (2002). Episodic memory: from brain to mind. Annu. Rev. Psychol. 53, 1–25. doi: 10.1146/annurev.psych.53.100901.135114
CrossRef Full Text | Google Scholar
Vandekerckhove, M., Bulnes, L. C., and Panksepp, J. (2014). The emergence of primary anoetic consciousness in episodic memory. Front. Behav. Neurosci. 7:210. doi: 10.3389/fnbeh.2013.00210
PubMed Abstract | CrossRef Full Text | Google Scholar
Vandekerckhove, M., and Panksepp, J. (2009). The flow of anoetic to noetic and autonoetic consciousness: a vision of unknowing (anoetic) and knowing (noetic) consciousness in the remembrance of things past and imagined futures. Conscious. Cogn. 18, 1018–1028. doi: 10.1016/j.concog.2009.08.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Vandekerckhove, M., and Panksepp, J. (2011). A neurocognitive theory of higher mental emergence: from anoetic affective experiences to noetic knowledge and autonoetic awareness. Neurosci. Biobehav. Rev. 35, 2017–2025. doi: 10.1016/j.neubiorev.2011.04.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Ward, L. M. (2011). The thalamic dynamic core theory of conscious experience. Conscious. Cogn. 20, 464–486. doi: 10.1016/j.concog.2011.01.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Watt, D. F., and Pincus, D. I. (2004). “Neural substrates of consciousness: implications for clinical psychiatry,” in Textbook of Psychiatry, ed. J. Panksepp (Hoboken, NJ: Wiley), 75–110.
Weiler, M., Northoff, G., Damasceno, B. P., and Balthazar, M. L. (2016). Self, cortical midline structures and the resting state: implications for Alzheimer’s disease. Neurosci. Biobehav. Rev. 68, 245–255. doi: 10.1016/j.neubiorev.2016.05.028
PubMed Abstract | CrossRef Full Text | Google Scholar
Withehead, A. N. (1929). Process and Reality. New York, NY: Macmillan.
Keywords: self, affect, emotion, archetype, consciousness, intentionality, subcortical midline structures (SCMSs), neurodynamic patterns